Балановская Е. Русский генофонд: свидетельства очевидцев
Каковы истоки русского генофонда? Какие племена и народы составили его основу?
Какие нашествия прошли, как волна над головой, не оставив и следа? Какие миграции - часто почти не отмеченные в летописной памяти - определили многие его современные черты?
На эти вопросы ищет ответы популяционная генетика, изучающая изменчивость генофонда в пространстве и времени.
КРАСКИ ДЛЯ ПОРТРЕТА
История формирования любого народа часто сложнее интриги приключенческого романа. Чтобы ее разгадать, нужно привлечь множество источников, каждый из которых говорит о той или иной стороне событий. И сейчас большие надежды возлагаются на генетику - ведь гены доносят до нас сведения о наших предках. Однако надежность свидетельств зависит от достоверности исходной информации. Решающее значение имеет спектр и количество популяций (Популяция — в данном контексте относительно обособленная группа населения, исторически сложившаяся на определенной территории и воспроизводящая себя в этих границах из поколения в поколение (прим. ред.), на основе изучения которых генетики делают выводы. За многие десятилетия тщательной работы антропологи, лингвисты, этнографы собрали детальные сведения практически обо всех народах мира. Огромный массив накоплен и биологическими дисциплинами — соматологией (Соматология — отрасль морфологии человека, изучающая вариации размеров и форм тела и его частей (прим. ред.), дерматоглификой Дерматоглифика — изучение деталей рельефа кожи (папиллярных линий) пальцев ладоней и стоп. Применяется в расоведении, криминалистике (прим. ред.), палеоантропологией.
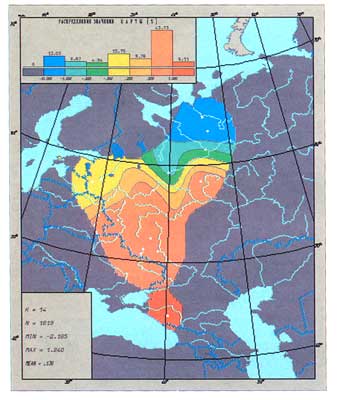
Первая главная компонента изменчивости русского генофонда
(по данным о частотах гаплогрупп Yхромосомы)
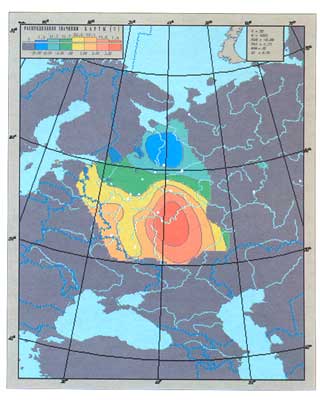
Первая главная компонента изменчивости русского генофонда
(по данным о классических маркерах)
На гистограмме указаны граничные значения, разделяющие шкалу изменчивости
признака на интервалы. Зона высоких значений признака окрашена в красно-коричневые цвета,
средних - в зеленые цвета, низких - в синие цвета
Еще недавно исследования генофонда протекали параллельными потоками. Слиянию мешало отсутствие технологии обобщенного анализа различных признаков, изученных к тому же в разных популяциях.
Сыграть объединяющую роль и провести синтез разнообразных данных о русском генофонде сумела геногеография. И сам термин, и понятие «генофонд», и идея связать процессы формирования народов с пространственным распространением генов принадлежат Александру Серебровскому (член-корреспондент АН СССР с 1933 г.), в 20-х годах XX в. писавшему в одной из своих работ: «...Современная география генов является результатом длительного исторического процесса, и когда мы научимся читать то, что записано в образах современного распространения генов, мы сможем прочитать подробную историю... человечества».
Сыграть объединяющую роль и провести синтез разнообразных данных о русском генофонде сумела геногеография. И сам термин, и понятие «генофонд», и идея связать процессы формирования народов с пространственным распространением генов принадлежат Александру Серебровскому (член-корреспондент АН СССР с 1933 г.), в 20-х годах XX в. писавшему в одной из своих работ: «...Современная география генов является результатом длительного исторического процесса, и когда мы научимся читать то, что записано в образах современного распространения генов, мы сможем прочитать подробную историю... человечества».
Генофонд — реальный объект. Не видимый ни в какие приборы, он имеет определенные физические параметры, структуру, занимает четко ограниченное пространство — ареал. Картографирование — единственный способ зримо представить этот объект. Поэтому создание компьютерных карт и их анализ — не дань моде, а необходимость и условие масштабного исследования. Вне картографической технологии невозможно ни описать географию сотен генов, ни тем более получить обобщенный «портрет» генофонда (а именно создание таких портретов авторы этой статьи рассматривают как одно из главных достижений представляемой ими лаборатории). Словом, геногеография не просто увеличивает количество соответствующей научной информации, а упорядочивает и преобразует ее, делая легко читаемой и доступной всем специалистам.
Правда, сегодня о генофонде, его разрушении и вырождении, средствах спасения чаше говорят не столько ученые, сколько общественные деятели и публицисты. Потому что исследователи могут позволить себе это, только располагая достоверными знаниями. И первый шаг к прогнозу будущего — обратиться к прошлому.
МЕТОДЫ ГЕНОГЕОГРАФИИ
Мы изучаем современное население, но сам анализ направлен на то, чтобы в ныне существующем генофонде различить черты истории его сложения. Именно поэтому в поле нашего интереса не урбанизированное или образовавшееся в результате недавних миграций население (тогда бы мы зондировали лишь недавнее прошлое), а коренное сельское (менее всего изменившееся с прежних веков). Именно поэтому мы ограничиваем рассмотрение «исконным», историческим ареалом русского народа, составляющим лишь часть современного. Русский народ формировался на территории, охватывающей центр Восточной Европы и ее Север. А определение «исконный» взято в кавычки не случайно: здешняя история дославянского населения на порядок дольше славянского.
Вместе с тем мы не говорим о специфически русских генофонде и генах. Ибо любая привязка биологического носителя наследственности к этносу неверна по сути — речь идет о разных системах координат: принадлежность к народу определяется самосознанием человека, генофонд же — концентрацией генов в определенном ареале. Как щепки в поток, гены через своих носителей — членов популяции — вовлекаются в исторический процесс, позволяя следить за его ходом века и тысячелетия. Возникает особая — историческая — связь генофонда и этноса.
Мы изучаем современное население, но сам анализ направлен на то, чтобы в ныне существующем генофонде различить черты истории его сложения. Именно поэтому в поле нашего интереса не урбанизированное или образовавшееся в результате недавних миграций население (тогда бы мы зондировали лишь недавнее прошлое), а коренное сельское (менее всего изменившееся с прежних веков). Именно поэтому мы ограничиваем рассмотрение «исконным», историческим ареалом русского народа, составляющим лишь часть современного. Русский народ формировался на территории, охватывающей центр Восточной Европы и ее Север. А определение «исконный» взято в кавычки не случайно: здешняя история дославянского населения на порядок дольше славянского.
Вместе с тем мы не говорим о специфически русских генофонде и генах. Ибо любая привязка биологического носителя наследственности к этносу неверна по сути — речь идет о разных системах координат: принадлежность к народу определяется самосознанием человека, генофонд же — концентрацией генов в определенном ареале. Как щепки в поток, гены через своих носителей — членов популяции — вовлекаются в исторический процесс, позволяя следить за его ходом века и тысячелетия. Возникает особая — историческая — связь генофонда и этноса.
Но жизнь исследователя слишком коротка в сравнении с жизнью популяции. Поэтому геногеография заменяет наблюдение во времени наблюдением в пространстве, а применяемые для этого инструменты — компьютерные карты — позволяют отслеживать одновременно микроэволюционные траектории множества генов. Чем больше статистических данных включено в подобную карту, тем детальнее будет восстановлена география исторического процесса в ареале генофонда. Конечно, границы популяции не представляют собой глухих заборов — сквозь них проходят потоки генов, однако на границах эти потоки не столь интенсивны, как в пределах собственного ареала популяции. Да и сами эти границы подвижны, текучи, хотя абсолютно реальны: их можно обнаружить и зафиксировать, например, по резким перепадам в частотах встречающихся генов, или изучая структуру миграций, связанных с заключением браков и созданием новых семей.
Даже имеющие общее происхождение генофонды под влиянием природных или демографических факторов от поколения к поколению удаляются друг от друга, что рано или поздно проявляется в антропологическом и генетическом своеобразии населения. Если отразить на картах изучаемые признаки, то окажется: в пространстве они распределены не хаотично. Увеличение и уменьшение частоты встречаемости генов происходит более или менее плавно, в результате чего не в отдельных географических точках, а на целых территориях они имеют сходные значения.
Наряду с картографической технологией важное место в геногеографии занимают банки данных. Дело в том, что объем сведений, используемых даже не в слишком масштабном исследовании такого рода, огромен, а сами исходные показатели обычно рассеяны во множестве статей. Благодаря своей структурной организации и запрограммированным функциям подобное хранилище становится еще и инструментом проверки, систематизации и анализа накопленных фактов. Поэтому прежде чем создавать атласы карт, пришлось создать банки данных «Русский генофонд», «Палеолит Северной Евразии», «Русские фамилии» и ряд других.
Перечисленные методы мы применяли при изучении адыгов, башкир, белорусов, марийцев, монголов, осетин, русских и представителей других народов. Наш сравнительный анализ показал: в генофонде населения Северной Евразии (включающей территорию бывшего СССР — европейскую часть России, Кавказ, Урал, Среднюю Азию, Казахстан, Сибирь и Дальний Восток) сохранена наибольшая часть мирового генетического разнообразия. Какие силы его поддерживают? Ведущим фактором, по нашим оценкам, стали многочисленные этносы, поэтапно («квантуемо») возникавшие в этих пространственных пределах на протяжении минувших тысячелетий.
ОТ ГЕНОВ - К ГЕНОФОНДУ
«Портрет» генофонда можно «нарисовать», лишь исследовав отдельные гены. Работа эта трудоемка, требует намного больших затрат времени и средств, чем, например, анализ антропологического состава населения. Чтобы изучить ДНК маркеры (а именно они привлекают сейчас внимание популяционных генетиков), надо выехать в экспедицию и в ходе ее провести обследование популяции. У выразивших согласие в нем участвовать берут образцы венозной крови. Причем только у индивидов, не связанных между собой кровным родством, к тому же их предки на протяжении двух поколений должны относиться и к данному народу, и к данной популяции. Такие пробы обычно берутся исключительно у мужчин — все маркеры и отцовской, и материнской линий наследования в этом случае оказываются представленными в одной выборке. Сохраняемые на холоде образцы крови срочно доставляют в мо-лекулярно-генетический центр для выделения ДНК, далее хранимой уже в морозильных камерах. Затем начинается следующий, самый интересный, но длительный и дорогостоящий этап исследования: определение у каждого индивида тех вариантов генов (точнее, вариантов ДНК), по которым одни популяции отличаются от других. В итоге выявляется их ДНК полиморфизм. Причем нельзя ограничиться одним или несколькими генами — для видения всей картины их палитра должна быть велика и разнообразна.
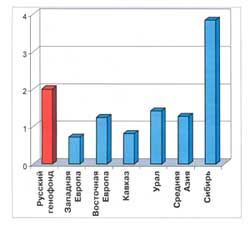
Гетерогенность русского народа в сравнении с типичной гетерогенностью народов регионов Евразии
Поясним подробнее. От каждого из родителей мы получаем по одному «комплекту» генов: один достается от отца, другой — от матери. Называются они аутосом-ными, и именно их у любого человека — абсолютное большинство. Однако есть и малые, но важные для изучения истории Homo sapiens исключения: речь идет о генах, переданных от одного из родителей и потому именуемых «однородительскими». Лишь мужчины и только от отца получают Ухромосому. У женщин ее нет. Зато от матери все мы — и мужчины, и женщины — получаем с яйцеклеткой особую ДНК, содержащуюся вне ядра — в митохондриях — и передающуюся в поколениях независимо от ядра. Гены человека можно представить как слова универсального языка наследственности. Тогда геном (или генотип) каждого из нас, включающий и аутосомные, и однородительские гены, сравним с составленным на этом языке неповторимым «текстом». А генофонд популяции, содержащий весь «словарный запас», — с совокупностью множества индивидуальных разнообразных «текстов».
Популяционная генетика занимается полиморфными генами, т.е. такими, которые встречаются не в одном, а в разных вариантах (аллелях) — «словах», различающихся лишь несколькими буквами. Каждый из вариантов-аллелей — результат мутаций (ошибок в написании «слов»), происшедших в далеком прошлом, но передающихся в цепи поколений до наших дней. Для изучения популяции важно, чтобы варианты не были очень редкими, а встречались в ней с частотой не менее 1 -5%. Впрочем, сколь бы ни было велико семейство аллелей, у индивида не может содержаться больше двух вариантов «слова» (два аллеля одного гена): по одному от матери и от отца. Если полученные аллели одинаковы, человек гомозиготен по данному гену, если различаются — гетерозиготен.
Аутосомные гены в процессе передачи рекомбиниру-ются («перетасовываются»). Так, если от отца вы получили полное собрание сочинений Федора Достоевского, а от матери — Агаты Кристи, то своему ребенку оставите случайно перетасованные тома — например, 1,2, 5, 8, 10-й Достоевского и 3, 4, 6, 7, 9-й Кристи. С одно-родительскими маркерами рекомбинации не происходят (поскольку получены только от одного из родителей) — они наследуются единым блоком и позволяют прослеживать историю материнских и отцовских линий. Такое их «полное собрание сочинений», целиком передающееся в ряду поколений, называют гаплотипа-ми митохондриальной ДНК (мтДНК) и Y хромосомы.
Суть всех генетических маркеров (физиологических, иммунологических, биохимических, аутосомных ДНК маркеров или же однородительских) едина: по результату тестирования (показываем ли участнику обследования книжку с цветными картинками, чтобы обнаружить дальтонизм, или проводим секвенирование ДНК, уточняя последовательность нуклеотидов), мы однозначно выявляем наличие или отсутствие у человека конкретных аллелей конкретного гена.
Подобная ситуация и с квазигенетическими маркерами: фамилиями, названиями рода. Хотя они и не «диктуются» генами (фамилия — явление языка и культуры, а не биологии), однако порой ведут себя подобно им и даже благодаря истории иногда оказываются в одной связке с генами.
ШИРОТНАЯ ИЗМЕНЧИВОСТЬ
Для выявления структуры русского генофонда мы проанализировали шесть массивов данных: два антропологических (соматология и дерматоглифика), два ДНК полиморфизма (мтДНК и Y хромосомы), еще один составили классические генетические маркеры (например, группы крови, гены ряда ферментов), а последний — география фамилий. Было важно выяснить, согласуются ли данные разных наук или противоречат друг другу, помогут ли они создать единый, целостный портрет? Каждый тип признаков — это «очевидец», рассказывающий о генофонде. И сравнение свидетельств помогает составить наиболее правдивый его образ.
Для выявления структуры русского генофонда мы проанализировали шесть массивов данных: два антропологических (соматология и дерматоглифика), два ДНК полиморфизма (мтДНК и Y хромосомы), еще один составили классические генетические маркеры (например, группы крови, гены ряда ферментов), а последний — география фамилий. Было важно выяснить, согласуются ли данные разных наук или противоречат друг другу, помогут ли они создать единый, целостный портрет? Каждый тип признаков — это «очевидец», рассказывающий о генофонде. И сравнение свидетельств помогает составить наиболее правдивый его образ.
Антропологические данные, на которые мы опирались, собраны в ходе двух масштабных экспедиций, проведенных под эгидой АН СССР в 50-х годах XX в. В этом наиболее крупном исследовании такого рода описан физический облик русского населения. Суммарно изучена 181 популяция по 18 признакам (длина тела, цвет глаз и волос, форма спинки носа, рост бороды и др.). География большинства этих характеристик довольно сложна. Простые закономерности обнаружены лишь для немногих признаков (скажем, для роста бороды: чем южнее популяция, тем средний рост бороды интенсивнее). В целом же антропологический облик, как следует из обобщенной карты так называемой канонической переменной (Карты канонических переменных и главных компонент выявляют «ведущие сценарии» изменчивости генофонда — то общее, что присутствует в большинстве карт отдельных признаков, но скрыто вуалью частной истории каждого признака (прим. авт.), меняется главным образом в направлении с севера на юг или обратно (ту же закономерность знают и лингвисты, выделяя северные, южные и смешанные среднерусские говоры). Но изменения происходят очень постепенно — ни о какой границе между севером и югом не может быть и речи. Это, скорее, главная ось, вдоль которой видна наибольшая изменчивость: двигаясь с севера на юг мы обнаружим намного большие различия, чем с запада на восток. Анализ признаков дерматоглифики также указал на широтную изменчивость, как на главную закономерность по второму массиву данных.
Массив так называемых классических генетических маркеров — возможно, самый важный: ведь в отличие от антропологических данных и фамилий это «настоящие гены», и по сравнению с недавно появившимися ДНК маркерами уже давно изучены во многих русских популяциях. Карта главной компоненты классических маркеров оказалась очень похожа на карту главной антропологической закономерности: снова постепенные изменения генофонда с севера на юг. Впрочем, ДНК маркеры не отстали от своих классических предшественников.
Из всего множества генетических маркеров Y хромосома - новейший и, по мнению многих ученых, наиболее многообещающий. Литературные сведения столь фрагментарны, что нам пришлось предпринять собственное исследование — собрать данные по 14 русским популяциям со всего «исконного» русского ареала и определить для них частоты гаплогрупп Y хромосомы (эту работу, как и по мтДНК, мы провели на базе Эстонского биоцентра, возглавляемого президентом Академии наук Эстонии Рихардом Виллемсом). Степень различий между популяциями (гетерогенность) по Y хромосоме оказалась гораздо выше, чем по классическим маркерам и мтДНК. Значит, для изучения русского генофонда маркеры Y хромосомы — наиболее мощный инструмент. Поэтому и для гаплогрупп Y хромосомы мы создали атлас карт их распространения в русском ареале. Оказалось, этот инструмент элегантно выявляет четкую и плавную широтную изменчивость русского генофонда: главный вектор, как стрелка компаса, вновь указывает то же направление — север-юг.
Согласие между всеми «очевидцами» не оставляет сомнений: выявлена реальная, объективная ведущая закономерность в структуре русского генофонда — широтная изменчивость.
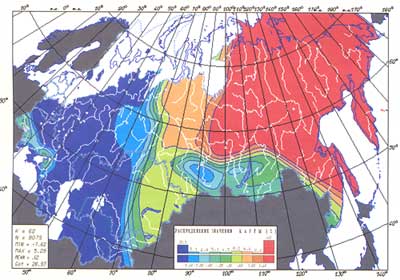
Первая главная компонента изменчивости материальной культуры верхнего палеолита на территории Северной Евразии
СЛАВЯНЕ И ИХ ПРЕДШЕСТВЕННИКИ
Эту закономерность обнаружили карты главных компонент. А вот на обобщенной карте другого типа — генетических расстояний — показано, насколько каждая из русских популяций по своему антропологическому облику отличается от среднестатистической. Похожие на среднюю встречаются повсюду и главным образом в центре русского ареала. Можно было бы ожидать, что «самые отличающиеся» располагаются на юге и севере. Однако популяции, значительно выпадающие из средних характеристик, образуют скопления, ядра по другой оси: одна группа на западе, другая — на востоке «исконного» ареала.
Для объяснения картины вспомним, как формировалось современное русское население. Славянские племена в течение нескольких веков продвигались на восток, колонизируя Восточно-Европейскую равнину и ассимилируя местные финно-угорские племена. В начале процесса славяне численно преобладали в западных областях, и эта доминанта заметна в облике населения «западных» ядер. В середине процесса смешение усилилось, особенно на территориях, являющихся ныне срединной частью русского ареала. В результате здесь образовался «среднестатистический» русский облик, составленный из славянского и финно-угорского компонентов. В конце же колонизации в восточных областях «исконного» русского ареала местное население численно преобладало над пришлым славянским, что неминуемо отразилось на внешнем облике: в «восточных» ядрах преобладают черты дославянского населения. Обо всем этом и свидетельствует наша гипотеза, подтвержденная тем, что зона «среднестатистического» русского населения соответствует продвижению границ Русского государства на восток в IX-XI вв., т.е. находится там, где происходило наибольшее смешение славян с финно-угорскими популяциями. К тому же западные «ядра» на антропологической карте совпадают с ареалами летописных славянских племен на археологической карте: каждое из зарегистрированных на западе ядер сопоставимо с племенем, упоминаемым в русских летописях (кривичи, вятичи, северяне).
ОСОБЕННОСТИ «ОЧЕВИДЦЕВ»
Выявив «главный сценарий» сложения генофонда, на втором этапе анализа надо всмотреться в своеобразие каждого используемого признака — будь то последовательности ДНК или черты внешнего облика, группы крови или фамилии. Может быть, особенности одного «очевидца» сообщат нам о генофонде то, что другие, в силу их особенностей, не откроют?
Так, среди дерматоглифических признаков особенно любопытно распространение европеоидно-монголо-идного комплекса — особого сочетания кожных узоров, хорошо дифференцирующих популяции запада и востока Евразии. Ожидание, будто у русского населения монголоидность возрастает к востоку или юго-востоку, не подтвердилось — в пределах «исконного» ареала этот комплекс распределен хаотично.
Казалось бы, у классических генных маркеров «особенностей» быть не должно. Но именно их «традиционность», то, что их изучают давно, придает им свойство, крайне важное для исследователей: по ним накопилось так много данных, что можно провести виды анализа, особенно требовательные к качеству и количеству исходной информации. Например, оценить такой параметр генофонда, как степень его внутренней гетерогенности (структурированности, дифференциации), т.е. узнать, насколько разные русские популяции отличаются друг от друга.
Выявив «главный сценарий» сложения генофонда, на втором этапе анализа надо всмотреться в своеобразие каждого используемого признака — будь то последовательности ДНК или черты внешнего облика, группы крови или фамилии. Может быть, особенности одного «очевидца» сообщат нам о генофонде то, что другие, в силу их особенностей, не откроют?
Так, среди дерматоглифических признаков особенно любопытно распространение европеоидно-монголо-идного комплекса — особого сочетания кожных узоров, хорошо дифференцирующих популяции запада и востока Евразии. Ожидание, будто у русского населения монголоидность возрастает к востоку или юго-востоку, не подтвердилось — в пределах «исконного» ареала этот комплекс распределен хаотично.
Казалось бы, у классических генных маркеров «особенностей» быть не должно. Но именно их «традиционность», то, что их изучают давно, придает им свойство, крайне важное для исследователей: по ним накопилось так много данных, что можно провести виды анализа, особенно требовательные к качеству и количеству исходной информации. Например, оценить такой параметр генофонда, как степень его внутренней гетерогенности (структурированности, дифференциации), т.е. узнать, насколько разные русские популяции отличаются друг от друга.
Чтобы ответить на этот вопрос, мы оценили показатель гетерогенности не только русского народа. Такой же анализ мы провели и для других этносов. Оказалось, все западно-европейские народы в целом гомогенны (популяции, например, французов генетически похожи друг на друга), а сибирские, напротив, гетерогенны (популяции, скажем, якутов очень различаются между собой). Промежуточное положение (умеренная гетерогенность) заняли народы Восточной Европы, Кавказа, Урала. На восточно-европейском фоне различия в русском генотипе очень велики. Они намного выше, чем средние генетические различия внутри каждого народа Западной Европы.
Митохондриальная ДНК как тип генетических маркеров сейчас наиболее популярна у исследователей во всем мире. Но данные по русским популяциям начали накапливаться только несколько лет назад. Поэтому мы пока располагаем надежными сведениями лишь по семи из них, и ограниченность информации не позволяет провести картографирование. А статистические методы выявляют, что по митохондриальной ДНК, как и по классическим маркерам, разные русские популяции весьма несхожи. Особенность митохондриальной ДНК — великое множество ее вариантов (гаплотипов), позволяющих сравнивать популяции по их «спектру». И по степени перекрывания спектров двух популяций можно судить об их родстве. Мы определили гаплоти-пы в русских популяциях и сравнили их со спектром у «соседей» по Европе. Оказалось, что русские наиболее похожи на остальные восточно-славянские популяции (30% «русских» гаплотипов встречены также у белорусов и украинцев). На втором месте по сходству стоят восточные финно-угорские народы (коми, удмурты, мари, мордва), на третьем — западные финно-угры (эстонцы, карелы, финны, саамы), затем идут западные славяне (поляки, чехи, словаки) и южные славяне (сербы, хорваты, болгары, боснийцы, словенцы). Итак, по гаплотипам мтДНК русский генофонд ближе скорее к финно-угорскому, чем к «праславянскому». Но самое главное, что как ранее по признакам антропологии, классическим генетическим маркерам и гаплогруппам Y хромосомы, мы по мтДНК вновь обнаружили: основной вектор изменчивости русского генофонда следует направлению «север-юг».
Самые необычные маркеры, на которые мы возлагаем большие надежды, — фамилии. По ним можно прогнозировать особенности генофонда там, где недостаточно времени и средств, чтобы изучать его напрямую, по генам. За несколько лет мы собрали данные о фамилиях около миллиона человек, представляющих сельское население в пределах «исконного» русского ареала. Пять регионов — Северный, Южный, Западный, Восточный, Центральный — стали каркасом исследования. К сожалению, у нас пока недостаточно информации по «стыкам» между ними. Поэтому картографирование провели только для 75 фамилий — по ним имелись данные для всего ареала. А закономерности остальных десятков тысяч фамилий изучены статистическими методами по «опорным» регионам.
Что же выяснилось? Почти каждая из 75 фамилий имеет свой географический ареал, за пределами которого она отсутствует или крайне редка. Даже такие, казалось бы, повсеместные фамилии — Иванов, Васильев, Смирнов — оказались не повсеместными: Ивановых очень мало на юге, Васильевы сосредоточены в северозападных областях, а Смирновы — в восточных и центральных. Впрочем, распространенность фамилий, как следует из наших обобщенных карт, подчиняется все той же широтной изменчивости, что и признаки антропологии и генетики, но эта закономерность затушевана — картографированный список недостаточно представляет многотысячный перечень русских фамилий.
Поэтому статистический анализ провели по всему спектру фамилий, а их мы обнаружили 65000.
По сложившейся практике, чтобы изучить коренное население, учли лишь деревни и небольшие города. Для отсева пришлых, «залетных» фамилий исключили встреченные в районной популяции менее чем у 5 человек. Оставшиеся 14 000 сочли условно коренными и работали дальше только с ними. Из них 250 оказались повсеместными: они встречаются в каждом из пяти упоминавшихся регионов, хотя и с разной частотой. Остальные рисуют своеобразный портрет каждого региона. В Западном преобладают «календарные» фамилии, т.е. произведенные от имен, упоминаемых в православном календаре. В Центральном наиболее характерны образованные от названий зверей, птиц, растений, в Восточном — приметные (Смирнов, Румянцев...). В Южном чаще всего встречаются профессиональные (Попов, Гончаров), а в Северном, хотя распространены календарные, очень много диалектных (Булыгин, Лешуков...). По частотам всех 14 000 фамилий регионы средней полосы (Западный, Восточный, Центральный) оказались похожи. А Северный и Южный отличаются и от нее, и друг от друга, опять напоминая о широтной изменчивости генофонда и стрелке компаса «север-юг».
Итак, рассмотрев основные результаты по типам признаков, характеризующих русский генофонд, констатируем: у них единый строй — широтное направление изменчивости.
В КРУГУ СОСЕДЕЙ
Выявление такого вектора тем удивительнее, что ожидать можно было как раз противоположного — направления изменчивости по долготе. Ведь для генофонда Восточной Европы (который включает русский как огромную составную часть) наши исследования показали: главная закономерность — изменчивость по долготе.
Как известно, русский народ сложился на основе восточно-славянских, балтских и финно-угорских племен, возможно, и тюркоязычных, ираноязычных -словом, едва ли не всех населяющих эту территорию этнических групп. Значит, именно долготная изменчивость должна была отразиться в нем, если он действительно складывался, как и Русское государство, «при-растанием», механическим включением в себя все новых групп населения. То, что выявлен иной — широтный — вектор, указывает на принципиальный факт своеобразия русского генофонда. Проявляется оно в том, что основное направление его изменчивости не сводимо к исходному, характерному для племен и народов, на основе которых он формировался. По-видимому, широтная изменчивость возникла или же усилилась в ходе собственной истории русского генофонда.
Выявление такого вектора тем удивительнее, что ожидать можно было как раз противоположного — направления изменчивости по долготе. Ведь для генофонда Восточной Европы (который включает русский как огромную составную часть) наши исследования показали: главная закономерность — изменчивость по долготе.
Как известно, русский народ сложился на основе восточно-славянских, балтских и финно-угорских племен, возможно, и тюркоязычных, ираноязычных -словом, едва ли не всех населяющих эту территорию этнических групп. Значит, именно долготная изменчивость должна была отразиться в нем, если он действительно складывался, как и Русское государство, «при-растанием», механическим включением в себя все новых групп населения. То, что выявлен иной — широтный — вектор, указывает на принципиальный факт своеобразия русского генофонда. Проявляется оно в том, что основное направление его изменчивости не сводимо к исходному, характерному для племен и народов, на основе которых он формировался. По-видимому, широтная изменчивость возникла или же усилилась в ходе собственной истории русского генофонда.
Отметим: долготный тренд (постепенное изменение значений в направлении «запад-восток») не ограничивается Восточной Европой. Это базовая, древняя закономерность генофонда всей Евразии. Проведенный нами картографический анализ археологических культур верхнего палеолита показал: 26-16 тыс. лет назад уже существовали резкие различия между населением Европы и Сибири. Однако в русском ареале, занимающем обширнейшую срединную часть Восточной Европы, на первый план вышла не эта фоновая изменчивость, а собственная, широтная. Впрочем, достоверно известно: в масштабе Евразии, разделяемой в первом приближении на западные, восточные и промежуточные области, русский генофонд относится к западному стволу.
В этой связи попробуем разобраться, какие последствия для него имело монголо-татарское иго — завоевание Руси в XIII в. и последовавшая зависимость от ханов Золотой орды. Конкретнее: каков вклад в наш генофонд степных завоевателей этого и нескольких последующих веков? Естественно предположить, что они оставляли своих потомков в местном населении, были и смешанные браки, и переселения отдельных групп — наверное, ни у кого не вызывает сомнения, что при политическом подчинении одного народа другому происходит смешение их генофондов. Но вот в какой степени?
Завоеватели-монголы происходили из степей Центральной Азии. Поэтому нам следует определить, насколько русский генофонд стал похож на генофонд монголов и их соседей (центрально-азиатский). Если, скажем генофонд поляков (славянского народа, который монгольские завоевания затронули лишь отчасти, а трехсотлетнее иго не коснулось вовсе) окажется не похож на центрально-азиатский, а русский в какой-то мере к нему приближен, то эта близость может свидетельствовать о реальном влиянии ига. Точнее, наличие такого сходства может быть следствием и более древних миграций, зато если сходства нет, то это однозначно укажет на отсутствие монгольского «следа завоевателей». Впрочем, речь идет о правдоподобной модели, ареальность, конечно, сложнее. Но популяционная генетика прибегает к моделированию, чтобы получить четкий количественный ответ.
По такому типу маркеров, как мтДНК, народы Центральной Азии и Европы различаются четко: в первой почти все население (более 90%) обладает восточно-евразийскими гаплогруппами, тогда как в Европе еще большая часть (свыше 95%) — иными, западно-евразийскими. Значит, процент восточно-евразийских гапло-групп мтДНК в русском генофонде прямо укажет на вклад центрально-азиатского населения. Фактически эта доля составила 2%, т.е. величину почти столь же малую, как и в генофонде поляков (1,5%) или французов (0,5%).
Не ошиблись ли мы? Все использованные данные, т.е. частоты гаплогрупп мтДНК, получены по большим выборкам разными исследователями и поэтому вполне достоверны. Может быть, войска Чингисхана и Батыя состояли не столько из населения Центральной Азии, сколько из степных народов Южной Сибири? Но и там восточно-евразийские гаплогруппы составляют ныне пусть не 100%, а только 60-80%, что несопоставимо больше 2%.
Еще одно возражение: мтДНК наследуется по материнской линии, а «генетический вклад завоевателей» мог быть скорее по отцовской. Результаты изучения гаплогрупп Y хромосомы («мужская» линия наследования) также не показывают значительной доли «степных» генов в русском генофонде.
Любопытны данные по эпикантусу (Эпикантус — складка у внутреннего уголка глаза человека, образованная кожей верхнего века и прикрывающая слезный бугорок. Характерен для монголоидной и некоторых групп негроидной расы (прим. ред.) — типичнейшему признаку монголоидной расы, в особенности характерному как раз для степного населения Центральной Азии. Антропологическое исследование, проведенное на огромной выборке русских (несколько десятков тысяч человек) практически не выявило случаев выраженного эпикантуса.
Итак, какой бы признак мы ни взяли, видим: русские — типичные европейцы, а азиатское завоевание оставило след в их истории, но не в генофонде.
Не беремся говорить о России и русских в культурном, историческом, гуманитарном смысле. Однако биологически их генофонд не является промежуточным между европейским и азиатским, он — типичный европейский. Уточним: самый восточный из этого ряда, стоящий «на передовой». Отдельные азиатские влияния прослеживаются в нем больше, чем в западных соседях. Но базовый, главный вывод, который следует из проведенного исследования — практически полное отсутствие в русском генофонде монголоидного вклада. Как нам кажется, последствия ига применительно к рассматриваемой проблеме заключаются не в закреплении генов завоевателей, а в оттоке русского населения, изменении направлений его миграций, в результате—и потоков генов, что, в свою очередь, сказалось на генофонде, в какой-то степени перестроив его. Возможно, влияние ига сказалось лишь в небольшой степени и только в восточной части ареала. Но не заметно, чтобы интенсивность смешений русского населения с пришедшим с востока, превышала обычный уровень в зоне контакта двух народов.
Доктор биологических наук Елена БАЛАНОВСКАЯ,
кандидат биологических наук Олег БАЛАНОВСКИЙ,
Лаборатория популяционной генетики человека Медико-генетического научного центра РАМН
кандидат биологических наук Олег БАЛАНОВСКИЙ,
Лаборатория популяционной генетики человека Медико-генетического научного центра РАМН
Иллюстрации авторов
"НАУКА В РОССИИ" № 2 (158) 2007




 Издательство «Свиньин и сыновья» выпустило несколько сотен самых разных по жанру, объему и авторам, но неизменно высококультурных изданий
Издательство «Свиньин и сыновья» выпустило несколько сотен самых разных по жанру, объему и авторам, но неизменно высококультурных изданий






